Discussion
Trypanosoma cruzi, as well as other trypanosomatids, lacks a heme biosynthetic pathway and therefore rely on the acquisition of this cofactor from their mammalian host or insect vector. Inside the insect vector,
T. cruzi may be in contact with hemoglobin (Hb) and heme derived from blood digestion. In vitro,
T. cruzi epimastigotes can use both hemin and Hb as a heme source, which are internalized via different pathways (
Lara et al., 2007).
TcHRG (
T. cruzi Heme Responsive Gene, previously named
TcHTE) is a protein belonging to the Heme Response Gene family. This protein is involved in heme uptake, probably being part of a heme transporter or regulating this activity. It is mainly expressed in the replicative life cycle stages of the parasites (epimastigotes and intracellular amastigotes). In epimastigotes,
TcHRG is localized to the flagellar pocket, where it is postulated that nutrients uptake takes place (
Merli et al., 2016). Our previous results allowed us to postulate a model proposing that
T. cruzi is able to sense the intracellular heme concentration and thus to regulate
TcHRG expression according to it (
Pagura et al., 2020). However, little is known about the use of Hb as a heme source, and conversely to
T. brucei and
Leishmania spp., no Hb receptor has been described yet in
T. cruzi.
In this work, we studied the role of
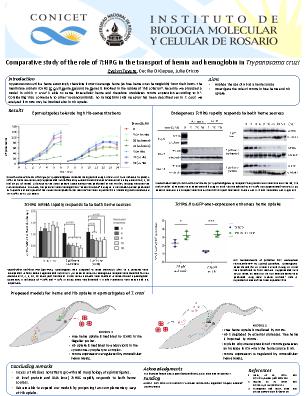
TcHRG in heme homeostasis when Hb is used as heme source. First, we analised the effect of different hemin and Hb concentrations on growth and morphology of Wild Type epimastigotes. When epimastigotes were incubated in a medium supplemented with high concentration of hemin, the growth and morphology were affected. However, they tolerated high Hb concentrations, exhibiting a typical morphology and no alterations in growth profile. Since we have polyclonal antibodies raised against
TcHRG, we were able to follow the endogenous protein by Western Blotting. Endogenous
TcHRG was detected as a strong signal in heme-starved parasites and became undetectable after the addition of hemin or Hb to the medium, confirming that
TcHRG responds also to Hb-derived heme. At mRNA level,
TcHRG was also higher in heme-starved parasites and lower when hemin or Hb was added to the medium. However, the response was less severe when Hb was used as heme source.
On the other hand, epimastigotes that over-express r
TcHRG.His-GFP incubated in Hb-supplemented medium have shown a significantly higher intracellular heme concentration compared to control epimastigotes. This finding was in agreement with previously reported assays using hemin as heme source (Merli
et al., 2016), suggesting that
TcHRG may also be involved in Hb import.
Based on these results, we were able to validate and expand our model by proposing two complementary ways of heme uptake in
T. cruzi. In the first one, Hb is endocytosed via the cytostome-cytopharinx complex and internally degraded. In the second one, Hb is externally degraded by parasite proteases and released heme enters the cell via
TcHRG. The second model would explain why over-expression of r
TcHRG.His-GFP produced an increase in the intracellular heme in Hb-supplemented parasites. In both models, once the parasite satisfies its heme requirements,
TcHRG expression (and therefore heme transport) decreases.