Authors
J Escalona1; J Bubis1; 1 Universidad Simón Bolívar, Venezuela Discussion
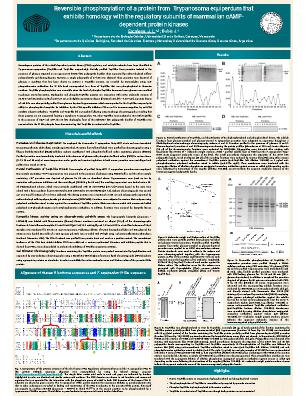
There is a deficit of knowledge regarding signaling pathways involving cAMP and cAMP-dependent protein kinase (PKA) in trypanosomes. In higher eukaryotic cells, the inactive PKA holoenzyme is a heterotetramer consisting of two regulatory (R) and two catalytic (C) subunits. Upon binding cAMP, the holoenzyme dissociates into an R subunit dimer containing four bound cAMP molecules and two free enzymatically active C subunits. Interestingly, proteins that show homology to the PKA R and C subunits have been identified in Trypanosoma equiperdum (TeqR-like and TeqC-like, respectively). T. equiperdum is the causative agent of dourine, a contagious venereally transmitted disease that affects horses and other Equidae. The TeqR-like protein differed from its mammalian counterparts given that it is monomeric, and is not capable of binding cyclic nucleotides, in spite of displaying two putative cyclic nucleotide binding (CNB) domains. Moreover, the kinase activity of TeqC-like was independent of cAMP, and was prompted upon nutritional stress driven by glucose fasting. Here, we evaluated whether the phosphorylation of TeqR-like by TeqC-like was feasible. Initially, we chromatographed the soluble fraction from T. equiperdum parasites isolated in the presence of glucose through a DEAE-Sephacel column. As shown by immunoblot analysis, the partially purified TeqR-like protein migrated as two polypeptide bands with apparent molecular masses of 55 kDa and 57 kDa. Anti-phospothreonine antibodies specifically recognized the band that migrated as a 57 kDa polypeptide, demonstrating that this band corresponded to the TeqR-like protein phosphorylated in threonine residues. Since glucose removal triggers a TeqC-like protein from T. equiperdum, the chromatographic separation was repeated using freshly purified parasites that were incubated in the absence of glucose. Only a band of 57 kDa that matched the migration of the phosphorylated polypeptide was obtained, and immunoblotting using anti-phosphothreonine antibodies confirmed that the phospho-TeqR-like protein was phosphorylated in threonine residues. TeqR-like phosphorylation was reversible since the level of phospho-TeqR-like decreased once glucose was readded to glucose starved-parasites. Gel filtration chromatography was carried out in order to determine the size and Stokes radius of the dephosphorylated and phosphorylated forms of TeqR-like. Dephospho- and phospho-TeqR-like proteins are monomers with native molecular masses of 54.93-57.41 kDa, Stokes radii of 3.42-3.37 nm, and slightly asymmetric shapes (frictional ratio f/fo = 1.36-1.32). A protein kinase of ~ 40 kDa was also partially purified from glucose deprived-trypanosomes, which corresponded to the TeqC-like enzyme by its ability to phosphorylate a specific substrate for PKA and PKA-like enzymes known as kemptide (sequence: LRRASLG), its inhibition by the PKA-specific heat-stable peptide inhibitor PKI-α and the synthetic peptide IP20 derived from PKI-α (sequence: TTYADFIASGRTGRRNAIHD), and its immunorecognition by anti-PKA catalytic subunit antibodies. TeqR-like and TeqC-like did not coelute following anion-exchange chromatography, revealing that these proteins are not associated forming a complex in T. equiperdum. Yet, when TeqR-like was incubated in vitro with TeqC-like in the presence of Mg2+ and ATP, the 55 kDa dephosphorylated form of the 55 kDa/57 kDa polypeptide doublet of TeqR-like was converted into the 57 kDa phosphorylated form, demonstrating that TeqR-like is a substrate for TeqC-like.